
Veterinary Clinics of North America: Small Animal Practice - Special Issue: Pediatrics - July, 1999
Congenital Deafness and Its Recognition
George M. Strain, PhD
Professor of Neuroscience, Veterinary Physiology, Pharmacology and Toxicology, School of Veterinary Medicine; and Associate Vice Chancellor, Office of Research and Graduate Studies, Louisiana State University, Baton Rouge, Louisiana, 70803
This work was supported by Grant No. 1R15DC01128-01 from the National Institutes of Health and by a grant from the American Kennel Club.
George M. Strain, PhD Comparative Biomedical Sciences School of Veterinary Medicine Louisiana State University Baton Rouge, LA 70803 (225) 578-9758 (225) 578-9895 (fax) strain@lsu.edu
Congenital deafness in dogs and cats is primarily of the hereditary sensorineural form associated with white pigmentation genes, although acquired forms of deafness are possible. Highest prevalence is seen in white cats, especially those with blue eyes, and the Dalmatian, with many other dog breeds affected to some extent. This deafness results from degeneration of the cochlear blood supply at age 3 to 4 weeks, presumably resulting from suppression of melanocytes by the white (cat) or merle or piebald (dog) genes. Mechanism of inheritance is not understood for most breeds. Such animals should not be bred and may present liabilities for their owners. Objective diagnosis of deafness, especially when unilateral, relies on the brain stem auditory evoked response, an electrodiagnostic test where electrical activity in response to a click stimulus is recorded from the scalp using needle electrodes and a special purpose computer. Client counseling guidelines are presented.
Sensory function in neonatal dogs and cats is primarily tactile, olfactory and gustatory. The visual and auditory senses, although partially functional at birth, exhibit significant postnatal development: in the dog, the eyes do not open until a puppy is 8 to 10 days of age, the ear canals do not open until it is 12 to 13 days of age, and mature system function up through the cortex is not present until it is 3 months of age or older.18,22 Similar delays are seen in the cat. As a result, disorders of these systems frequently escape early detection.
Deafness can be described as (1) congenital or late onset, (2) hereditary or acquired, and (3) conductive or sensorineural.14,15 The most commonly occurring deafness forms in dogs and cats are congenital hereditary sensorineural deafness, late onset acquired sensorineural deafness, and late onset acquired conductive deafness. Distinguishing between hereditary and acquired deafness is generally not possible without breeding trials, although an assumption of hereditary deafness can be made in breeds with a high prevalence of deafness. The most common form of deafness in young dogs and cats is congenital hereditary sensorineural deafness, with acquired conductive and acquired sensorineural appearing on rare occasions.
PATHOPHYSIOLOGY OF DEAFNESS
Perception of sound first requires transmission through the outer and middle ears to the cochlea for transduction by neural hair cells. Perception results from (1) transmission of transduced auditory information from the cochlea by the eighth cranial nerve to the dorsal and ventral cochlear nuclei, the inferior colliculus, the medial geniculate nucleus of the thalamus, and the primary and secondary cortical auditory areas on the temporal lobe; and (2) attention to the arriving information. Commingling of ipsilateral and contralateral auditory information occurs at multiple steps in the ascent up the auditory pathway. As a result, unilateral hearing loss rarely results from lesions or disease affecting auditory structures above the eighth nerve. Central deafness (unilateral or bilateral) in the absence of severe neurologic disease is clinically unknown in veterinary medicine and is not considered further here.
Conduction Deafness
Conduction deafness results from blockage of sound transmission to the cochlea as a consequence of occlusion of the ear canal or middle ear cavity, or from developmental defects. Occlusion may result from excess cerumen production, from otitis externa or media, or from foreign objects. Developmental defects, which are uncommon, may include atresia of the tympanum or ossicles, fusion of the ossicles, or collapse of the ear canal from cartilaginous weakness or incomplete development. Conduction deafness may be partial or complete and may be reduced by intervention in some cases. Clearance by the body of the mucopurulent discharge and detritus from otitis media may require weeks to months after termination of the infection; hence, recovery of auditory function is delayed. Hereditary forms of conduction deafness have not been identified in domestic species, but the appearance of such a disorder from a spontaneous genetic defect is possible.
Congenital Acquired Sensorineural Deafness
Congenital acquired sensorineural deafness, which is uncommon, can result from in utero or perinatal exposure to ototoxic compounds such as maternal treatment with aminoglycoside antibiotics,21 in utero or perinatal otitis or meningitis, anoxia, or even trauma. Breeders with animals belonging to breeds with a high prevalence of hereditary sensorineural deafness may suggest acquired causes of deafness rather than confront the breeding and other implications of the presence of a hereditary disorder. Sensorineural deafness, whether hereditary or acquired, is the consequence of cochlear hair cell loss through primary or secondary mechanisms (see below).
Congenital Hereditary Sensorineural Deafness
Congenital hereditary sensorineural deafness is usually seen in dog and cat breeds with white pigmentation. In the small number of canine breeds where it is not associated with white pigmentation (Doberman and other dog breeds not carrying piebald or merle genes),23 deafness results from the type of hair cell loss that is a primary event with unknown cause. In breeds of dogs carrying the piebald or merle genes and breeds of cats carrying the white gene, the hair cell loss is secondary to degeneration of the cochlear blood supply. Figure 1 shows a cross section of one turn of the cochlea, demonstrating the separation of the cochlea into three parallel ducts: the scala vestibuli, the scala media (or cochlear duct), and the scala tympani which joins at the apex of the cochlea with the scala vestibuli. The outer margin of the scala media is covered by a vascular bed, the stria vascularis. The stria is responsible for secretion of endocochlear fluid and maintenance of its high K+ concentration which is essential to sound transduction by the sensory hair cells. In pigment-associated hereditary deafness, this vascular bed degenerates, resulting in secondary loss of hair cells and deafness. The cause for the strial degeneration is unknown, but histologic studies have demonstrated an absence of strial melanocytes, whose presence or postnatal development is suppressed by the piebald or merle genes. The function of melanocytes in the stria is unknown, but they appear to be critical to maintenance of elevated K+ levels in the scala media and survival of the stria. Whether hair cell death is from primary or secondary mechanisms, the loss is permanent, as mammals are unable to regenerate cochlear neuronal tissue.
In the Dalmatian, postnatal auditory function development has been shown to proceed normally up until 3 weeks of age, at which point the strial degeneration produces rapid loss of hair cell function.6 A similar time course is likely in other breeds of dogs and cats with pigment-associated deafness, but it has not been studied. Likewise, the time course of congenital deafness in canine breeds not associated with white pigmentation has not been documented, but it is likely that deafness is present at birth or shortly thereafter. As a consequence of the documented 3 to 4 weeks of age at which time deafness appears, hearing testing, as described below, is typically not performed until an animal reaches at least 5 weeks of age.
A wide variety of breeds of dogs have been reported to have congenital deafness (see Table 1); not all of these cases of deafness have been shown to result from hereditary causes. The breeds for which the prevalence is known to be high are highlighted in bold, although similar high rates may occur in other breeds that do not yet routinely receive testing. Prevalence rates measured by the author are shown in Table 2 for the canine breeds most often presented for hearing testing services. The highest rates are seen in Dalmatians, of which 30% are deaf in one or both ears; rates in other strongly affected breeds range from 8% to 20%. Typically, there are two to three unilaterally deaf animals for every bilaterally deaf animal. In breeds with white versus non-white phenotypes (Bull Terrier, English Cocker Spaniel), there is a clear increased prevalence in the white phenotype. Prevalence rates for pure feline breeds have not been measured, but are highest for the breeds carrying the white gene (Table 3), especially in cats with blue eyes. Deafness in 256 mixed breed white cats was reported as 12% unilateral and 38% bilateral, for a total of 50% of cats being affected (reviewed in Delack3). The prevalence of deafness increases as the number of blue eyes increases from zero to two, but not all blue-eyed white cats are deaf. Deafness prevalence (unilateral and bilateral) in mixed breed white cats was 17%, 40%, and 85% for zero, one, or two blue eyes, respectively.9
GENETICS OF DEAFNESS
Pigment-associated deafness has been reported since the last century,1,5,8-10,12 but the hereditary mechanisms are not yet fully understood. The merle gene, which is responsible for the pattern of dark and light hair in breeds such as the Collie and Shetland Sheepdog, is a simple autosomal dominant gene.7 Dogs that are homozygous for the merle gene are usually deaf and frequently solid white, blind, and sterile. Heterozygotes have an increasing tendency to be deaf as the percent of white in the coat increases. Although the merle geneis a dominant gene, the deafness associated with it is not inherited as a simple dominant (or recessive) disorder.
The piebald and extreme piebald genes, which are responsible for white hair in nonmerle breeds of dogs, are simple autosomal recessive genes.7 As a result, in breeds with major white areas on the body, the animals are homozygous. An example is the Dalmatian, all of which are breed are homozygous for the extreme piebald gene. The underlying coat color of black (dominant) or liver (recessive) is covered with white by the extreme piebald gene, and spots are produced through the white by the dominant ticking gene. The lightness or heaviness of the spotting is thus controlled by the ticking gene and not the extreme piebald gene, and it is not a factor in the prevalence of deafness.19 Based on studies of the Dalmatian, deafness in breeds carrying the piebald genes is neither simple recessive nor dominant. Breeding of bilaterally hearing parents routinely produces deaf offspring; thus, the mechanism is not simple dominant. Breeding of two bilaterally deaf parents produces both deaf and hearing offspring; the latter would not occur if the defect were simple recessive and both parents were homozygous. There is no evidence for X-linked or mitochondrial hereditary mechanisms. As a consequence, inheritance of deafness associated with the piebald genes must be polygenic or must involve incomplete expression or penetrance, or some combination. Molecular genetic studies of deafness disorders in humans and mice suggest that this deafness may result from a defect in a gene responsible for regulation of the piebald genes such as one of the homeobox class of genes. Evidence for this comes from further findings in Dalmatians. A patch in Dalmatians is a large black or liver area present at birth when the puppy is otherwise solid white; patches are disallowed in the Dalmatian breed standard. The patch appears to result from weak expression of the extreme piebald gene, resulting in a failure to cover the underlying coat color; patched Dalmatians are statistically less likely to be deaf than unpatched animals.19 When the extreme pieblad gene is strongly expressed, brown pigment is suppressed in the iris, resulting in blue eyes (and frequently an absence of pigment in the tapetum lucidum), and melanocytes are suppressed in the cochlear stria vascularis, resulting in deafness. Blue-eyed Dalmatians are statistically more likely to be deaf than brown-eyed animals.19 The blue eye is permitted in the breed standard in the United States but not in Canada, Mexico, or Europe. The prevalence of deafness is lower in Europe (combined unilateral and bilateral deafness is 21% in the United Kingdom and 18% in Holland compared to 30% in the United States), and breeding away from blue eyes was shown to reduce deafness in Norway.4 It is not known if differences exist for deafness associated with the two different piebald genes.
Deafness in the Doberman Pinscher, which is accompanied by transient vestibular dysfunction, is transmitted by a simple recessive mechanism.23
Pigment-associated congenital hereditary sensorineural deafness in the cat is linked to the white gene, which is dominant over color and is unrelated to albinism.13 On occasion these cats will have a head spot and usually have one or two blue eyes. Although the white gene is dominant, not all carriers are deaf; thus, deafness is not simply inherited.
BEHAVIORAL INDICATORS OF DEAFNESS
Newborn puppies and kittens with undeveloped auditory and visual function use other sensory cues for their feeding, elimination, and locomotion behaviors. As auditory development proceeds, they can detect loud noises, despite the unopened ear canal. Breeders relying on this for home testing may find themselves to have been in error at a later date. Behavioral testing of hearing after opening of the canal relies on detection of a response to sound stimuli in the absence of other detectable sensory signals. These noises should be produced outside of the visual fields, avoiding visual cues, vibratory cues, touch, and air movements. Behavioral testing has limited value; animal responses rapidly adapt even when hearing is present, stressed animals with intact hearing may fail to respond, and unilateral deafness cannot be detected. In unilaterally deaf animals, the only behavioral sign of deafness is a difficulty in localizing the source of a sound, and many animals adapt to that also. Behavioral deafness detection with young animals in the home is difficult, as the deaf young cue off the behavior of their littermates. A puppy or kitten that does not awaken in response to a loud noise is almost certainly bilaterally deaf, but the unilaterally deaf cannot be detected with any reliability. As a consequence, behavioral hearing assessment of animals in the clinic or home is of limited reliability, and electrodiagnostic tests are used for objective assessment.
ELECTRODIAGNOSIS OF DEAFNESS
The most widely used electrodiagnostic test of hearing is the brain stem auditory evoked response (BAER), also known as the brain stem auditory evoked potential (BAEP) or the auditory brain stem response (ABR). This test was first used in veterinary research applications in the 1970s, and in clinical veterinary applications in the early 1980s. The BAER detects electrical activity in the cochlea and auditory pathways in the brain in much the same way that an electrocardiogram detects electrical activity in the heart.17 The response waveform consists of a series of peaks labeled with Roman numerals: peak I is produced by the cochlea and auditory nerve, and later peaks are produced within the brain. The response from an ear that is deaf is an essentially flat line. In the sample recordings in Figure 2, the Dalmatian puppy in tracing A could hear in the tested ear, although the Dalmatian puppy in tracing B was deaf in the tested ear, with an essentially flat response. Because the response amplitude is quite small, it is necessary to average the responses to multiple stimuli (clicks) to unmask them from the other unrelated electrical activity that is also present on the scalp (e.g., electroencephalographic activity, muscle activity).
The response is collected with a special computer through small subdermal needle electrodes: one is placed in front of each ear, one is placed at the top of the head, and a ground electrode is placed either between and behind the eyes or on the neck. It is rare for a dog to show any evidence of pain from the placement of the electrodes - if anything the dog objects to the gentle restraint and the presence of wires hanging in front of its face. A stimulus click (air-conducted) produced by the computer is directed into the ear with a foam insert earphone. Each ear is tested individually, and the test usually is complete in 10 to15 minutes. Sedation or anesthesia is unnecessary unless the dog becomes extremely agitated, which can usually be avoided with patient and gentle handling. Sedation or anesthesia does not materially affect the BAER.
The click stimulus used contains most of the audible frequencies of the dog and cat, with the exception of the very highest perceived frequencies. Accordingly, the BAER is a frequency nonspecific test that is more useful for detecting the presence or total absence of auditory function without quantifying hearing loss in decibels. Assessment of the normalcy of a response is based on identification of the presence of Peak I within a narrow expected time frame (which varies based on the equipment used) and the presence of the expected pattern of peaks. With progressive hearing loss, there is a reduction in the amplitude of the BAER peaks and an increase in peak latencies; thus, a subjective assessment of partial hearing loss can be made but not quantified, and differing degrees of loss in different frequency ranges cannot be determined. Diagnosis of partial hearing loss based on the BAER is done only with great caution, as a number of technical factors can affect peak amplitude and latency in subjects with normal hearing.17 Fortunately, partial hearing loss is rare in puppies or kittens.
The BAER demonstrates maturational changes.22 Because the greater portion of the BAER originates in the brain stem, there is less postnatal development than is seen in tests of other sensory modalities; however, postnatal development is greater in altricial species like the dog and cat than in precocial species like the horse and cow. Full maturation of the BAER occurs by 40 days in the dog and the cat. The BAER can be recorded in response to loud stimuli prior to the opening of the ear canal, but this is not of use as it predates the age at which deafness is manifested.
In some circumstances, it is useful to be able to differentiate between sensorineural and conductive deafness, as this can affect breeding decisions and whether a young animal is placed in a show home or a pet home. When a BAER indicates deafness in an animal in which conduction deafness might be suspected (i.e., long-eared breed, recent ear infection), the test is repeated with a mechanical transducer that transmits the stimulus click as a vibration through bone rather than through air conduction.20 Because the cochlea is imbedded in bone, the bone-conducted BAER bypasses the outer and middle ears, the sites of conduction blockade, and directly activates the cochlea. The response appearance is the same as an air-conducted BAER, but the peaks occur at a shorter latency due to the shorter path traversed by the stimulus (Figure 2).
A limited availability of BAER testing sites blocks some potential users from access, but the number of test locations is increasing beyond the original veterinary school sites. The equipment cost of approximately $20,000 and an absence of formal veterinary training programs outside of neurology residencies have impeded ready access. A listing of available national and international sites is maintained at the author's Web site,16 which is also a resource of additional information on deafness.
CLIENT COUNSELING ISSUES
Advice to clients faced with a deaf puppy or kitten varries based on breed, animal age, home environment, unilateral versus bilateral deafness, and other factors. Bilaterally deaf animals present a variety of liabilities and emotional land mines, more so for dogs than for cats. Deaf animals are at risk of injury or death from undetected dangers such as motor vehicles. When startled, they may reflexly bite, which is a special concern around infants and toddlers. Anxious or aggressive personalities may develop in deaf dogs from constantly being subjected to startle, and familiar family members and friends may be attacked without warning or cause. Not all deaf dogs develop these problems, and no data exist on prevalence rates for such events, but there is no way to predict which animals may or may not have these experiences. Based on inherent temperament differences, there may also be variation between breeds in the likelihood of such problems. Bilaterally deaf dogs are very difficult to raise and train; as a result, they often end up in animal control shelters. From there, they may be reclaimed by breed rescue groups, after which the cycle may begin again. House cats present fewer problems, but outdoor cats may fall victim to motor vehicles. An emotional cost is invariably paid by the owners of those animals that cannot cope or adjust to their disability, both from the perspective of management and that of facing the decision as to whether to euthanize an animal with which an emotional attachment has formed. Unilateral deafness does not pose such problems.
Because of the many problems associated with bilaterally deaf dogs and the surplus of available puppies, the Dalmatian Club of America has an official position calling for the euthanasia of deaf puppies, with the emphasis on breeders rather than new owners. Similar sentiment is held by the official organizations of other breeds with high deafness prevalence rates, but these groups have not adopted such an official policy. This position has generated considerable controversy within the purebred dog community, the general dog-owning community, the veterinary community, and the human deaf community, but it is probably appropriate when divorced from emotional considerations.
Resources exist for those owners opting to keep a deaf dog. Books have been written on living with a deaf dog,2 including directions for training deaf dogs to respond to American Sign Language signs, and web pages have been posted with information and support content. Deaf dogs and cats learn to respond to flashing porch lights and vibrating collars,16 and can cue off the behavior of other animals in the household. In such circumstances, the owner should be instructed in protecting the animal from the inherent dangers associated with deafness: both dangers to the deaf animal and dangers to people around the deaf animal.
Genetic counseling for owners of deaf dogs and cats cannot be presented with total assurance because of the incomplete knowledge of mechanisms of deafness inheritance. It can be stated that deaf animals in breeds with a high known prevalence of either unilateral or bilateral deafness should not be bred; unilaterally deaf animals have the genetic defect but have one ear spared. Over the long run, such breedings have the probability of producing more deaf animals. In addition, it may not be advisable to breed to animals from litters which had a high percentage of deaf animals or from lines with a history of producing high percentages of deaf animals. The most conservative approach avoids any possible introduction of defective genes, but may be difficult in certain breeds. A deaf animal from one of these breeds must be assumed to have hereditary deafness instead of acquired deafness unless the clinical history convincingly indicates otherwise. Because of the association between deafness and blue eyes in the Dalmatian, it is also advisable to not breed blue-eyed dogs from those breeds in which the blue eye is not a standard part of the breed phenotype.
If a deaf animal is presented from a breed with no history of notable numbers of deaf animals, the guidelines for advice are murkier. The most conservative approach is not to breed any affected animal, especially if the breed is one carrying piebald or merle genes, unless the clinical history suggests a likely acquired cause such as otitis or drug ototoxicity. Even these latter cases are not without risk unless precedent documentation of normal hearing exists.
In an effort to promote research to reduce deafness and provide data for potential breeders, several national breed organizations in this country have set up hearing registries either managed by the breed organization (e.g., English Setter Association of America) or by a contracted second party (e.g., Dalmatian Club of America managed by the Institute for Genetic Disease Control in Animals, Bull Terrier Club of America managed by the Orthopedic Foundation of America). The first registry is closed, although the second is open and the third offers the option of being either closed or open to qualified members of the public. Current canine deafness research focuses on determining mechanisms of inheritance from extended pedigrees and on identifying the responsible defective genes using molecular biological techniques.
References
1. Anderson H, Henricson B, Lundquist P-G, et al: Genetic hearing impairment in the Dalmatian dog. Acta Oto-Laryngol Suppl 23:1, 1968
2. Becker, SC: Living With a Deaf Dog. Cincinnati, Susan Cope Becker, 1998
3. Delack JB: Hereditary deafness in the white cat. Compend Cont Ed Pract Vet 6:609, 1984
4. Greibrokk T: Hereditary deafness in the Dalmatian: Relationship to eye and coat color. J Amer Anim Hosp Assn 30:170, 1994
5. Hudson WR, Ruben RJ: Hereditary deafness in the Dalmatian dog. Arch Otolaryngol 75:213, 1962
6. Johnsson LG, Hawkins JE Jr, Muraski AA, et al: Vascular anatomy and pathology of the cochlea in Dalmatian dogs. In: de Lorenzo, AJD (ed): Vascular Disorders and Hearing Defects. Baltimore: University Park Press, 1973, p 249
7. Little CC: The Inheritance of Coat Color In Dogs. New York: Howell, 1957
8. Lurie MH: The membranous labyrinth in the congenitally deaf collie and Dalmatian dog. Laryngoscope 58:279, 1948
9. Mair IWS: Hereditary deafness in the white cat. Acta Otolaryngol Suppl 314:1, 1973
10. Mair IWS: Hereditary deafness in the Dalmatian dog. Arch Otorhinolaryngol 212:1, 1976
11. Pujol R, Hilding D: Anatomy and physiology of the onset of auditory function. Acta Otolaryngol 76:1, 1973
12. Rawitz B: Gehörorgan und Gehirn eines Weissen Hundes mit blauen Augen. Morphol Arbeiten 6:545, 1896
13. Searle AG: Comparative Genetics of Coat Color in Mammals. London: Logos Press, 1968
14. Strain GM: Congenital deafness in dogs and cats. Compend Cont Ed Pract Vet 13:245, 1991.
15. Strain GM: Aetiology, prevalence, and diagnosis of deafness in dogs and cats. Brit Vet J 152:17, 1996
16. Strain GM: Deafness in dogs and cats. Located at: http://www.lsu.edu/deafness/deaf.htm
17. Strain GM: Electrophysiological assessment of auditory function. Proc. 15th ACVIM Forum 15:617. 1997.
18. Strain GM, Jackson RM, Tedford BL: Postnatal development of the visual-evoked potential in dogs. Am J Vet Res 52:231, 1991
19. Strain GM, Kearney MT, Gignac IJ, et al: Brainstem auditory evoked potential assessment of congenital deafness in Dalmatians: associations with phenotypic markers. J Vet Internal Med 6:175, 1992
20. Strain GM, Green KD, Twedt AC, et al: Brain stem auditory evoked potentials from bone stimulation in dogs. Am J Vet Res 54:1817, 1993
21. Strain GM, Tedford L, Jackson RM: Postnatal development of the brainstem auditory-evoked potential in dogs. Am J Vet Res 52:410, 1994
22. Strain GM, Merchant SR, Neer TM, et al: Ototoxicity assessment of a gentamicin sulfate otic preparation in dogs. Am J Vet Res 56:532, 1995
23. Wilkes MK, Palmer AC: Congenital deafness and vestibular deficit in the doberman. J Small Anim Pract 33:218, 1992
November 15, 1998
Figure 1. Cross-section of the cochlea. The organ of Corti rests on the basilar membrane, with its hair cell cilia embedded in the tectorial membrane. The stria vascularis on the outer margin of the scala media secretes the endocochlear fluid of the scala media and maintains a high K+ concentration essential to sound transduction by the hair cells. Sensorineural deafness can result from primary or secondary loss of cochlear hair cells. (From Bloom W, Fawcett DW: A Textbook of Histology, ed 10. Philadelphia: WB Saunders, 1975; with permission.)
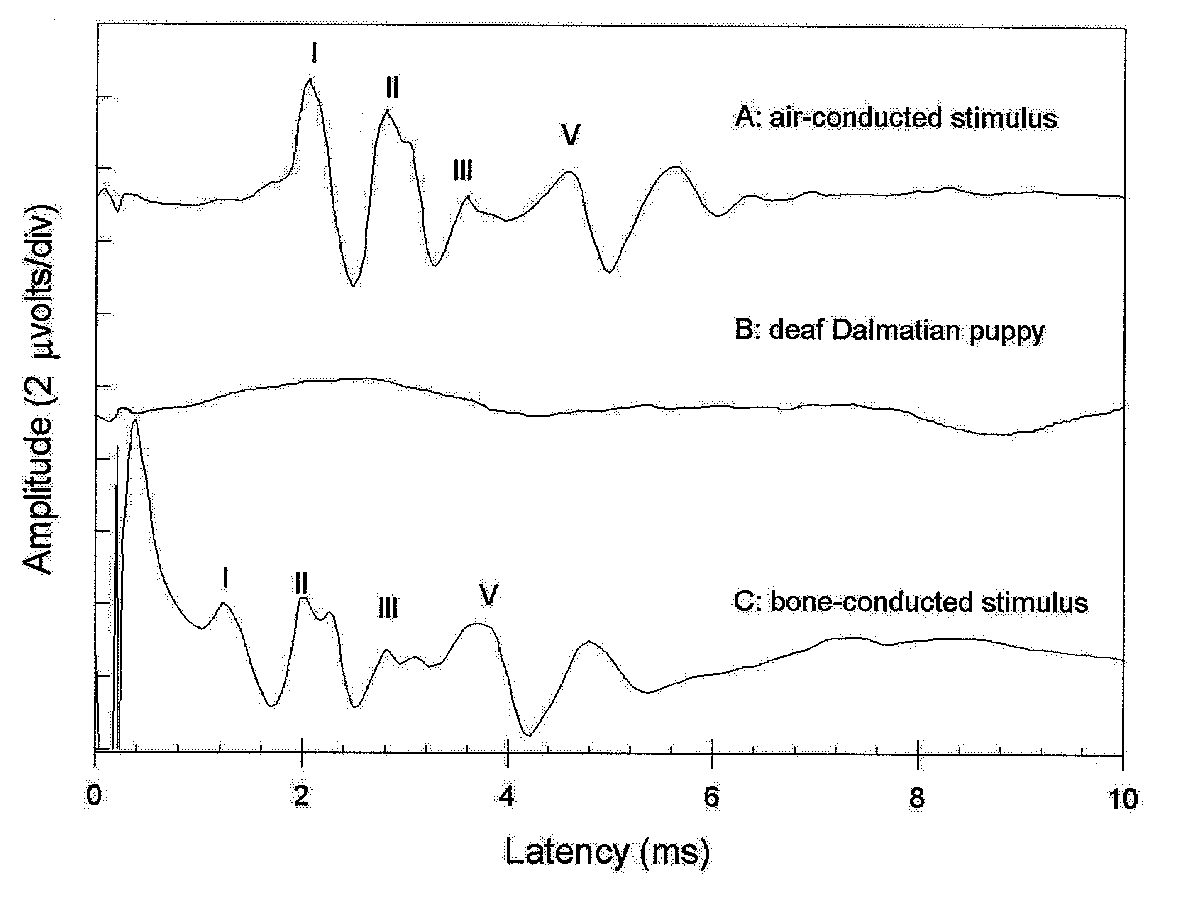
Figure 2. Brain stem auditory evoked responses (BAER) recorded from puppies; cat BAER appear similar. A. A BAER from a normal dog in response to an air-conducted click stimulus. Peak I in the response is generated by the cochlea and VIIIth cranial nerve, while later peaks are generated in brain stem structures. B. A BAER from a dea0f Dalmatian. C. A BAER from a normal dog in response to a bone-conducted click stimulus.